ПРОМО АКЦІЯ
Ставка без Ризику
Робіть ставки без ризику на вибрані матчі! Виграйте або поверніть 100% ставки, якщо програєте!
Букмекерська контора Mostbet на ринку Росії, Казахстану, України, Узбекистану, Азербайджану з 2009 року належить StarBet NV. Працює у 92 країнах світу, за ліцензією №8048/JAZ2016-065, онлайн, через офіційний сайт. Наземних точок у закладі немає. На сайті є два розділи: букмекерська контора та казино.
Mostbet
Ефективність ставок у Мостбет середня, маржа 10, у каталозі доступні події з 24+ видів спорту, відповідно у будь-якій грі виплата може становити 95%. На футбол у вищих лігах приймаються 85 видів ставок як гарантія того, що будь-який гравець зможе отримати свою 95% виплату:
Оцінка 5/5 користувачами:
Мінімальна сума депозиту складає 10 $. Поповнення здійснюється практично вмить після створення заявки в особистому кабінеті на сайті букмекерської контори. Для поповнення/виведення коштів доступні платіжні системи:
Висновок здійснюється через особистий кабінет – “Висновок з рахунку” – вказати спосіб, номер рахунку – “Замовити платіж”. Гроші прийдуть на реквізити, які краще використати раніше для поповнення балансу. Термін залежить від типу платіжної системи, від кількох хвилин до 72 годин.
Важливо! Перед виведенням проставте суму свого поповнення за ставками, вказавши коефіцієнт 1,3 або грайте на ці гроші в казино в будь-які слоти, віртуальний спорт або лайв казино.
Букмекерська контора орієнтована на футбол, сайт відстежує всі значні події – матчі, чемпіонати, локальні зустрічі, прем’єр-лігу.
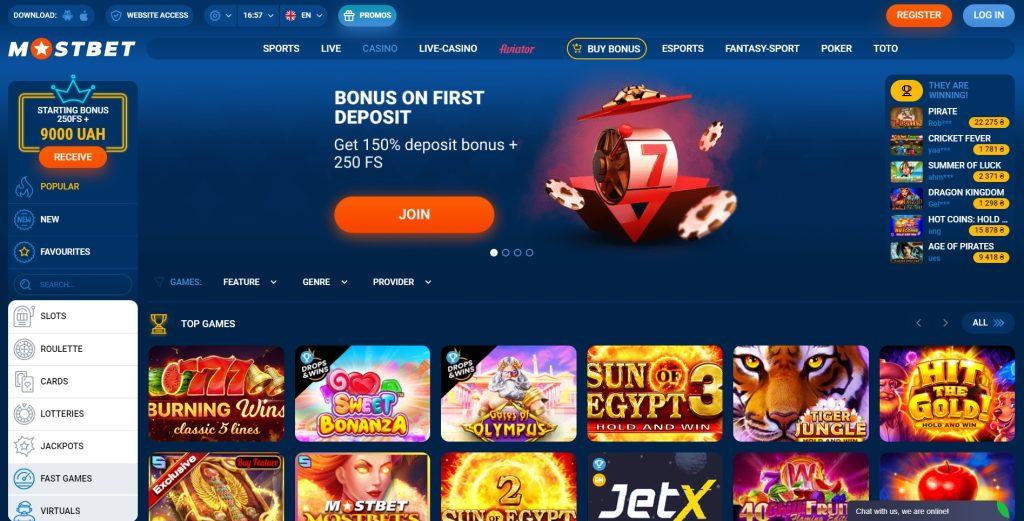
Інтерфейс сайту Мостбет зручний, у верхній частині над логотипом компанії знаходиться меню налаштувань та інші функціональні вкладки:
![]()
Основну частину сторінки займає таблиця зі спортивними подіями, ліворуч під смугою з іконками приховано бічне меню. Крім розділів «Головна» та «Майбутні» (показує останні спортивні події в часі), «Футбол» з 6 підрозділами:
Ви можете дивитися матчі країнами, вибираючи зі списку з назвами. Розділ “Футбол”/”Клуби” – розташований над таблицею з подіями, можна вибрати “ПРЯМИЙ ЕФІР” та “Лінія” – для звуження пошуку. Або за назвою команди – типу змагання через рядок пошуку.
![]()
Додатки приховані в меню під значком «Ще»:
Ви можете стежити за змаганнями команд у CS:GO та інших іграх, у букмекерській конторі є віртуальні види спорту (футбол, регбі, волейбол, понад 10 різновидів).
Праворуч знаходиться букмекерський купон, внизу сторінки інформаційні вкладки (Про нас, правила, контакти, мобільна версія, партнерка, FAQ – з відповідями на популярні питання). Нижче розташовані логотипи платіжних систем, назви популярних футбольних клубів та кнопка «Завантажити» для завантаження мобільної версії сайту.
Сайт ліцензований, завдяки сучасній системі шифрування дані каппера знаходяться у безпеці, поза досяжністю третіх осіб.
Ви можете зробити ставку на:
Робіть прогнози про те, хто переможе, а хто зможе відігратися і скільки очок. Важливо уникати згадування армії та битви на війні, подібні теми негативно впливають на рейтинг закладу, адже він діє легально.
Mostbet пропонує безліч бонусів для новачків та постійних гравців. Вітальний бонус допомагає гостям букмекерської контори швидше освоїтися:
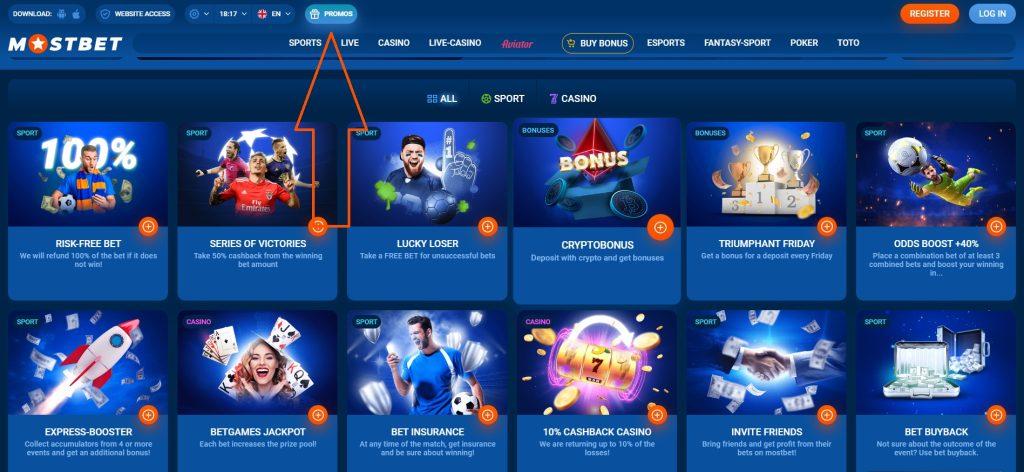
Кешбек у казино також становить 10%. БК, як і раніше, регулярно проводить акції в дати знаменних свят, щоб бути в курсі новин, необхідно зареєструватися і відразу підписатися на розсилку. Активні гравці мають доступ до ставки без ризику страхування безкоштовної ставки. Дарують подарунки на день народження.
Щоб скористатися всіма пропонованими бонусами, вам необхідно зареєструватися на платформі та зробити поповнення балансу будь-яким доступним на Мостбет способом. Новачкам видаються «плюшки» за 1-4 депозити, є обмеження за мінімальною сумою. Після виконання умов подарунки будуть нараховані автоматично.
| Бренд | Мостбет |
| Офіційний сайт | Mostbet.com |
| Люстерко | Мостбет Украина |
| дата заснування | 2009 рік |
| Ліцензія | Курасао |
| Власник | StarBet NV через CMS Trust NV |
| Типи ігор | Ставки на спорт, ставки в реальному часі, казино, живе казино, авіатор, кіберспорт, покер |
| Кількість ігор | 3700+ |
| Поле | 6-7% |
| Мови | RU, EN, UZ, AZ, TR, HI, IR, UA, KZ, ES, EN, FR, UR, HY, AG, BR, BY, CZ, PL, RO, PE, BD, NO, PT, CL |
| Країни | увесь світ |
| Заборонені гео | США, Франція, Великобританія, Іспанія та Італія |
| Платіжні системи | Visa, Mastercard, Maestro, СВІТ, Visa Qiwi Wallet, Neteller, Piastrix, Webmoney, Skinpay, ecoPayz; Білайн, Мегафон, Теле2 та МТС, Bitcoin, Litecoin, Ripple та Ethereum |
| Валюти | AED, ALL, AMD, AUD, AZN, BDT, BND, BRL, BYN, CLP, CNY, COP, CZK, DKK, EUR, GBP, GEL, GHS, HRK, HUF, IDR, ILS, INR, IRR, JPY, KES, KHR, KRW, KZT, mBT, MKD, MNT, MXN, MYR, NGN, NOK, PEN, PHP, PKR, PLN, RON, RSD, RUB, RWF, SEK, SGD, THB, TWD, TZS, UAH, UGX, USD, UZS, VND, XAF, XOF, ZAR |
| мін. депозит | 5$ |
| мін. виплата | 10 $ |
| Місячний ліміт зняття | 2000 $ |
| Бездепозитний бонус | Ні |
| Бонус на депозит | 500 $ + 250 ФС |
| гроші | h60 |
| Підтримувати | [email protected] |
| Платформа казино | Власний |
Букмекерська контора стане зручним майданчиком для новачків, які бажають освоїтись у світі ставок та вибудувати свою стратегію без великих фінансових втрат. Мінімальна ставка одна і поширюється на всю систему – 10 $. Максимальна – змінюється під впливом факторів:
Наприклад, на ФІФА, Барселону чи серію матчів НБА у плей-офф іноді ставлять 1 000 000 доларів, і це не межа. Місцеві події менш актуальні.
Зареєструватися на сайті можна 4 способами, натиснувши кнопку «Реєстрація» у верхньому правому кутку. Відкриється форма, де гість вибере відповідний варіант:
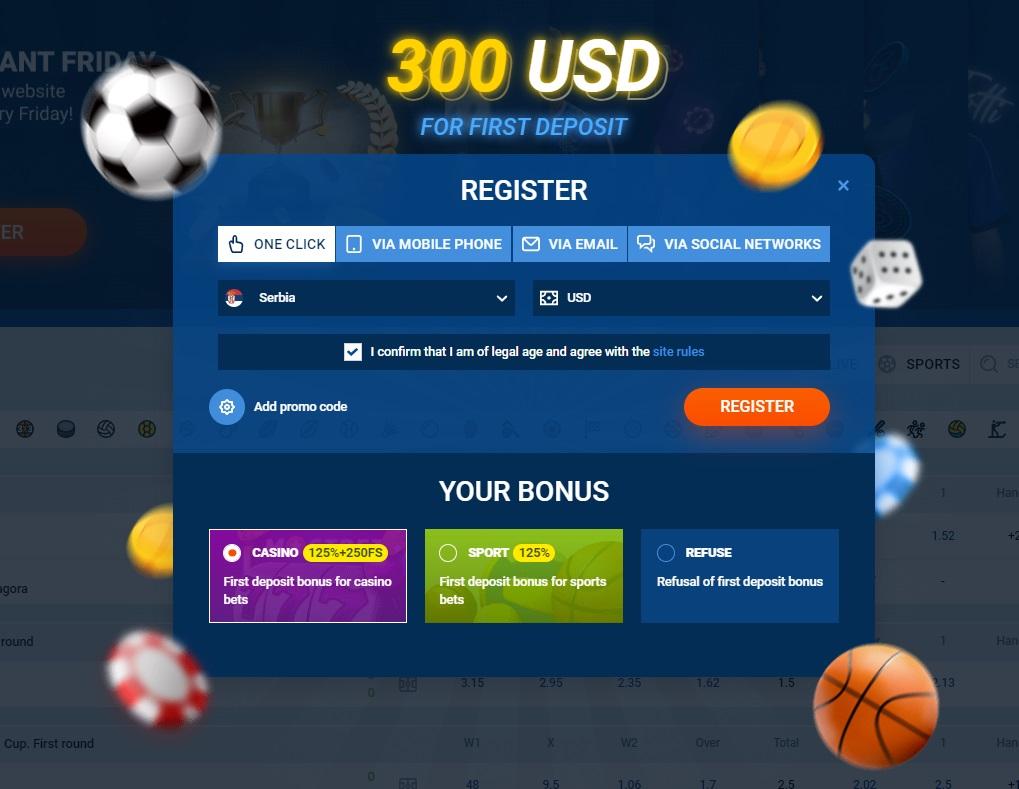
Букмекерська контора в рамках правил безпеки вимагатиме від нового гравця підтвердити свою особу, надавши йому відскановані копії сторінок паспорта/посвідчення водія та сторінки електронного гаманця/боків банківської картки. Всі дані захищені сучасними протоколами – SSL та TLS, що запобігає ризику фішингу та інших порушень.
Необхідно підтвердити номер телефону/адресу поштової скриньки/соціальну мережу, на неї прийде перевірочна SMS з кодом, її необхідно ввести протягом найближчих 72 годин після реєстрації.
На сайті ви можете завантажити мобільну версію платформи для пристроїв iOS та Android. Функціонал практично ідентичний браузерній версії, доступні такі опції:
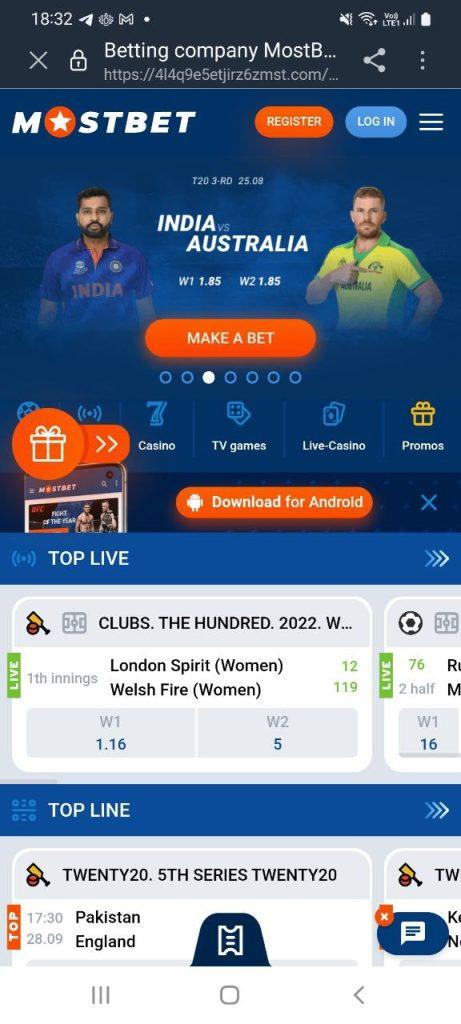
Мінус – доступні не всі відеотрансляції, середня оптимізація, залежить від технічних характеристик пристрою, немає результатів матчів, можна дивитися епізоди і прев’ю. Пошук через пошукові фільтри. Доступні швидкі програми, авторизуватися можна під своїм обліковим записом.
Клієнти Mostbet можуть поставити будь-яке питання про роботу букмекерської контори або казино у службу підтримки, вибравши будь-який із доступних способів зв’язку:
Найшвидша відповідь – чат. Спілкування можна почати зі звичайного – “Вітаю” і чекаєте на відповідь, можете одразу поставити запитання. Оператори завжди на місці, відповідають протягом 1-2 хвилин в залежності від завантаженості сайту. Доступно 24/7.
Оцінивши функціонал та інші аспекти роботи Мостбету, можна виділити переваги сервісу:
Mostbet має Telegram-канал, де ви можете отримувати останні новини про рейтинги, трансфери та гравців. Майже всі значні події пов’язані зі світом спорту. Сподобалися трансфери гравців Пердю, Лапорт, Жоан, Жузес, Гарман, Ювентус. Анонси перемог у матчах, таймах, парі-матчах, прес-релізи, ФІФА, півзахисники, матчі – топ та локальні, зіграні на перемогу чи нічию, Прем’єр-ліга. Статті виходять 1-2 на день регулярно.
На головній сторінці сайту є вкладка «Казино», натиснувши яку можна перейти в однойменний гральний заклад. У каталозі представлені слоти від Playson, Spinomenal, Pragmatic Play та інших ліцензованих операторів. Стартовий бонус для новачків 250 фриспінів + 500 $. крім слотів є рулетка та карткові ігри (баккара, покер, блекджек) та живі ігри з живими дилерами. Авторизація не потрібна, букмекерська контора та казино знаходяться на одному сайті.
Казино Mostbet пропонує своїм користувачам більші можливості. На сайті в каталозі представлено безліч онлайн-слотів та захоплюючих гральних автоматів на будь-який смак. Є класичні із фруктами, ковбойські чи сучасні із оновленою механікою, наприклад, авіатор. Більше того, поряд із різними азартними казино-розвагами користувачам доступні і звичайні онлайн-стратегії, в яких можна заробити реальні гроші. Загалом, способів розважитись і отримати чималу нагороду достатньо, головне вчасно зупинитися та забрати виграш.
Основний напрямок казино – ігрові автомати. Є потрійні та п’ятірні барабани з різними символами на тему древнього Єгипту, космічної одіссеї, детектива. Щоб вибрати відповідний слот для гри, відкрийте відповідну сторінку сайту та почніть свою захоплюючу подорож гральними джунглями. Основна мета ігрових автоматів – отримання прибутку внаслідок збігу 3-х символів на одній із встановлених ліній. Кількість останніх може змінюватись від 9 до 200 і більше. Чим їх більше, тим вищий виграш, але нижче коефіцієнт. Казино “Мостбет” славиться своєю доступністю. У неї можуть грати всі, навіть громадяни з невеликим статком. У слотах можна робити ставки від 10 $, а максимальний виграш за ставку становить 30 000 $. Окрім основного виграшу, у деяких слотах розігрується джекпот, сума якого може його перевищувати.
Щоб отримати дозу адреналіну, не обов’язково одразу грати на реальні гроші. По-перше, радимо вам використовувати демо-режим у слотах. Його наявність у тій чи іншій грі залежить від провайдера, який надав його ігровому закладу. У більшості слотів він є. У демо-режимі користувач грає не на реальні гроші, а на віртуальні. Для користувача створюється спеціальний ігровий обліковий запис з фальшивою валютою. Відвідувач грального закладу не поповнює рахунок, а відразу починає робити ставки в слотах і крутити барабани або грати в інші ігри на свій вибір. Якщо він «зіллє» усі гроші, то на гаманець це ніяк не вплине. Щоб знову почати грати, просто перезавантажте сторінку у браузері.
Для гри в демо-версію казино Мостбет не потребує реєстрації. Вам просто потрібно відкрити цікавий слот і дочекатися його завантаження. Все дуже просто. Демонстраційний режим – це свого роду тренувальний режим.
Користувач опрацьовує стратегію, вивчає особливості гри на тому чи іншому автоматі, вибудовує свою статистику так, щоб при поповненні ігрового рахунку одразу почати заробляти, а не зливати свій капітал. Тренувальний режим дає гравцям безліч переваг:
Головною перевагою гри на віртуальні гроші є можливість отримати адреналін без ризику свого гаманця.
Незважаючи на наявність демо-режиму, багато користувачів не можуть жити без адреналіну, а отримати його більше можна лише через реальний ризик. Тому одразу грають на реальні гроші. Ризик – ключ на щастя, але для таких людей не має значення, виграли вони чи ні, адже навіть після великого провалу вони продовжать переходити на сторінку гральних автоматів та відкриють їх у платному режимі.
Але як грати на гроші у казино Мостбет? Все дуже просто. Вам необхідно поповнити ігровий рахунок та вибрати будь-який слот. Зрозуміло, зробити це можна лише після реєстрації, яка триватиме лише кілька хвилин. Нагадуємо, що онлайн-казино дарує новачкам вітальний бонус. Щоб використовувати його, вам потрібно буде ввести промо-код IGRA150.
| Назва казино | Мостбет |
| Офіційний сайт | www.mostbet.com |
| Мобільний додаток | Андроїд | IOS |
| Назва компанії | Бізбон Н.В. |
| Ліцензія | ні. 8048/ЯЗ2016-065 |
| Фундамент | 2009-09-09 |
| Підтримувати | Живий чат; [email protected] |
Турнір – це чудова можливість отримати високий додатковий прибуток, адже адміністрація казино завжди добре заохочує активних користувачів. Суть гри полягає у протистоянні з іншими користувачами, які також хочуть отримати додатковий виграш. Фонд турнірів завжди дуже великий. Іноді його вартість сягає неймовірних 2-3 мільйонів доларів. Основні турніри:
Інформація про майбутній турнір завжди представлена на головному екрані сайту онлайн-казино. Також інформація про нові турніри розсилається на телефони та пошту, щоб активні гравці не пропустили унікальну пропозицію стати мільйонером. До особливостей турнірних битв можна віднести те, що навіть користувач, що програв, отримує виграш.
Мінімальна сума нарахування від 15 доларів, а максимальна іноді перевищує 3 тисячі. Сума нарахування залежить від активності користувача. Чим довше і частіше він робить ставки, тим вищим він піднімається в рейтингу. Головне завдання – піднятися якомога вище в турнірній таблиці. Плюс бонус, отриманий у результаті турнірної гри, зараховується на рахунок для гри та виведення. Це реальні гроші, які гравець може вивести на карту або витратити в іншій букмекерській грі.
Приємним подарунком для клієнтів грального онлайн-закладу стане бонусна програма у Most Bet Casino. Бонуси та акції надаються будь-яким казино, але їх обсяг індивідуальний. Хтось нараховує більше бонусних грошей на окремий рахунок, а інший онлайн-заклад дає більше фриспінів. Яка бонусна програма у цьому закладі?
Ні для кого не секрет, що будь-який бонус – це насамперед спосіб залучити нових відвідувачів до вашого закладу. І, як показує практика, цей метод працює дуже добре. Іноді достатньо вказати в описі, що кожен отримає 200% на рахунок, як одразу реєструється, але при цьому бонус не можна використовувати в жодних слотах. Адміністрація казино прописує конкретні умови, коли та як можна відіграти бонус.
До особливостей даного ігрового закладу можна віднести те, що бонуси використовуються не лише як маркетингова програма, а й як спосіб добре заробити. Гральний заклад пропонує своїм користувачам безліч приємних подарунків:
Гральний клуб «Мостбет» регулярно пропонує своїм користувачам приємні бонуси. Так, наприклад, зараз є ще одна пропозиція. Щоб скористатися вітальним бонусом, вам знадобиться промокод “IGRA150”, він надасть вам 100% та +250FS.. Його необхідно ввести під час реєстрації у спеціальне поле. Подивіться на сторінку створення нового облікового запису, вона зазвичай знаходиться внизу сторінки. Насправді, ви можете отримати до 125% на свій ігровий рахунок, тому що коефіцієнт збільшення залежить від суми внесених коштів. Чим більше ви переведете, тим вищим буде загальний дохід від бонусної акції.
Грати на мостбет це круто. Особливо в компанії з друзями. Зібраись разом і грати на мостбет це топ.
Официальный сайт мостбет я очень часто играю в игры и класное казино и мне бывает везет всем советую пробуйте супер.
Всім рекомендую завантажити додаток на ios, там дуже зручно. Мені подобається
Сайт мені дуже заходить з його зручністю, багато функцій і багато ігор.
Грати на мостбет одне задоволення. Такої кількості ігор я ніде ще не бачив. Ні на одному онлайн казино – тільки мостбет





